戻る
さまざまなクローン蚕
1 クローン蚕
1996 年に羊の体細胞核由来のクローン個体(ドリー)が,イギリス・ロスリン研究所キャンベルによってつくりだされ,高泌乳牛のクローン個体作出などに応用されている。クローンは同じ個体を起源として,これとまったく同じ遺伝子型を持った個体の群をいう。高等動物においてはイギリスのゴードンが 1962 年に一匹のカエルの細胞核をあらかじめ核を除いておいた受精卵に移植して,遺伝的に均一な個体群を作り出すことに成功したのが手はじめである。
カイコでは単為発生によってこのような個体をつくりだす試みは早くからなされ,ロシアのチコミロフ( 1885 )がその先駆者である。未受精卵が自然に発生を始め,幼虫が孵化する現象はしばしば観察されるが,その発生頻度は低く,多数の単為発生個体を得ることはできなかった。この発生率を格段に高めたのは,やはりロシアのアスタウロフの功績である。
2 メスからメス(雌性単為発生)
写真をクリックするとカイコの単為発生の実験の
手順が見られます
写真提供東京農工大学、蜷木理・横山岳 |
アブラムシは夏の間中,単為生殖を営み,メス親からメスの子だけ生まれることはよく知られた現象である。鱗翅目昆虫でも,ミノムシにはこのような産雌現象をくり返す種があることが知られている。
アスタウロフの方法は,未交尾のメス蛾の腹部を切開して卵管を取り出し,ガーゼの袋に入れ水に浸して指先でもみほぐし,卵粒をバラバラにする。この卵を 46 ℃の温湯に 18 分間浸漬した後,常温の水で5〜 10 分冷やしてから乾燥して, 15 〜 16 ℃の部屋に 3 日間置く。この実験では,成功率が 50 〜 60 %と高かったが,育ったカイコはメスばかりであった。アスタウロフはこの原因を卵母細胞中の減数分裂をする前の核が,温度処理を受けたため,減数分裂を行うことができず,直ちに体細胞分裂に入ったためであろうと考えた。もしそうならば,単為発生個体では,遺伝子構成が母親個体とまったく同じであることが期待されるが,アスタウロフは,これを標識遺伝子を用いた実験で証明した。
3 一代雑種を固定する
アスタウロフはこの方法で, F 1 から始めて11代目まで,毎代単為発生を行わせることができた。しかし残念なことに,第二次世界大戦に際会して,ドイツ軍がモスクワに侵入したため,この系統はことごとく失われてしまった。
このようにして単為発生をくり返した場合,虫の強健度や繭の形質はどうなるだろうか。蚕糸科学研究所に勤務する東京農工大学卒業の大隈琢己は,アスタウロフの実験を追試し,単為発生を3代継続してくり返した。その結果, F 1 を出発点として,単為発生を3代くり返した場合,単為発生蚕の各世代の示す繭形質の大きさはほぼ同じレベルに保たれ, F 1 を 100 とした場合,全繭重では三世代の平均 82. 6 ,繭層重では 71. 0 となり,単為発生世代の方が F 1 より明らかに 20 〜 30 %下回った。これは単為発生を行わせるために,卵に高温処理を行ったことに原因しているものと思われる。
4 二精子メロゴニー(雄性単為発生)
橋本春雄は産下直後の卵を高温処理する一連の実験の中で,卵核がまったく発生に関与しないで,精核だけが発育したとみられる例外型が多数出現してくることをみつけた。これらの個体は異常なく発育し,生殖力にも異常が認められなかったので,ハプロイド(単数体)とは考えられない。種々の点から,この個体はおそらく精核2個が合体して生じた,二精子メロゴニーらしいと考えられた。そこで橋本は正常型のメスに標識遺伝子を持つオスを交配しこの推定を確かめた。カイコでは受精に際し,一個の卵に数個の精子が侵入する多精現象が普遍的にみられることは川口栄作により1926年観察され,ひろく認められているところである。
5 ホモ雄をつくる
イネやタバコでは花粉の培養から半数体の植物を得て,それを倍加して二倍体の育種親として使うようになっている。育種の観点からいえば,ホモ型オスを高い確率で得る方法が望まれた。
スツルニコフはその効率を高める方法の開発に成功した。このためにはまず雌原核を不活化することが必要で,これには低温(マイナス10℃)処理が有効なことをテルスカヤが発見していた。そこでこの方法を利用して,その手順をつぎのように示した。
①まずアスタウロフの方法により卵巣から卵を取り出す。
②水洗,乾燥後18〜20℃に15〜20時間保護する。
③卵を取り出して,その後マイナス11℃に30分置く。
④冷却した卵は16℃に移し,三日間保護する。
⑤人工ふ化処理を希望する場合はここで行う。
なお,この実験をいっそう効果的に行いたい場合には,単為発生を起こしやすい系統を利用するとよいようである。この実験ではメス親には優性遺伝子を,オス親には劣性遺伝子をマーカーにつけた系統を使う。マーカーとしては Pe , re などの卵色遺伝子, ch などの蟻色, od , ok などの油蚕遺伝子,血液色に関する y などが利用できる。
6 倍数体
蚕はもちろん,高等生物では体細胞のもつ染色体数は両親の生殖細胞(卵子および精子)から受けとった染色体数の和,すなわち 2n である。ところが自然界でもときとして, 3n とか 4n ……といった染色体数をもつ個体があらわれてくることがあるし,また人為的にも誘発することができる。
多倍数体はその出現によって遺伝情報量を一挙に増加できるので,生物進化の数あるしくみの中でも重要な要因と考えられている。また一般に植物の多倍数体は二倍体にくらべると,細胞が大きく,作物などでは体も巨大化して,利用価値のすぐれたものが多い。したがってカイコでも多倍体をつくって,大きな繭をつくらせ,生糸の収量を増加させることはできないかという考えが,研究者たちの頭に浮かんだ。今から 50 〜 60 年前のことである。
カイコで倍数体の誘発を最初に報告したのは,田中義麿・川口栄作( 1931 )による遠心力を用いた実験である。その後,川口は大規模な実験を行って, 1936 年その結果を詳細に報告した。
この実験では産下後1〜3時間の卵を遠心機にかけ,毎分 3500 回転で 10 分間処理した。処理後ふ化した蚕を育て,合計 17193 匹を調査した。このうち少なくとも三倍体である可能性をもつもの 846 匹について倍数性別に分けてみると, 183 匹( 21.6 %)は四倍体, 572 匹( 67.7 %)は三倍体, 91 匹( 10.7 %)は二倍体であった。なお,これらのうち一匹のメスは F 2 の性状から六倍体であることが判明した。
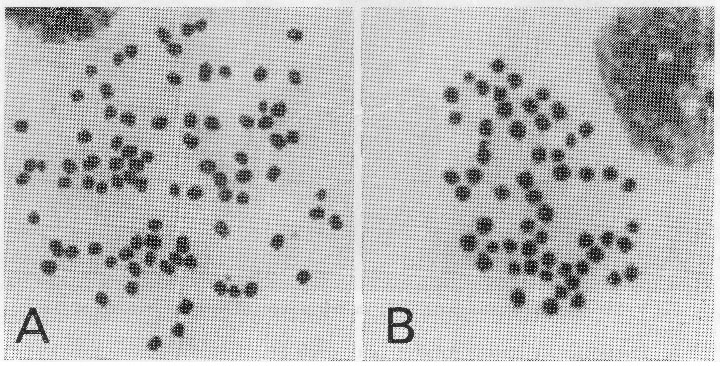
A:3倍体蚕の染色体(3n=84)、B:2倍体蚕の染色体(2n=56)<拡大> |
オスについては細胞学的に倍数性を確かめたが,メスは産下した卵の大きさや形状から倍数性を推定した。すなわち四倍体は産下する卵が大型で,そろっていること,二倍体オスとの交雑の子が三倍体であることなどから判定がついた。
川口についで,橋本は産下直後の卵に高温処理(40℃,1時間)を行う方法で,やはり多数の三倍体や四倍体を得ることができた。その後,広部達道はコルヒチンの希薄溶液( 0.05 〜 0.4 %)を産下2時間半以内の卵に塗布する方法で,二倍体と多倍体の混在する混数体を得たし,また橋本は遺伝的モザイク系統から四倍体がかなりの頻度で出現することを監察報告している。このようにカイコでは倍数体を誘発することは比較的容易である。
7 倍数体の利用
四倍体では産下卵の大きさが二倍体にくらべて明らかに大きい。その他カイコの倍数体についてはつぎの測定結果がある。多倍体は二倍体にくらべて細胞の形は大きいが,絹糸腺細胞数や産下卵数などはかえって多倍体の方が小さくなっている。結局,カイコでは多倍体には経済的な有利性は認められないと結論された。
しかも多倍体とくに三倍体では,生殖細胞形成にあたり染色体の対合が乱れて,精子や卵子に配分される染色体数が不均等となり,高い不妊性を示すので,これが利用にあたって大きな障害と考えられた。これに対し,四倍体メスでは減数分裂における染色体対合は比較的正常だが,それでも二,三の染色体については異常対合像がみられる。川口の得た四倍体ではオスはすべて生殖力を欠いていた。これらのことから,わが国では 1940 年代以降,倍数性の利用は顧みられなくなった。
8 四倍体のクローン
日本の蚕遺伝学者たちは倍数体の研究に見切りをつけてしまったが,ソ連ではアスタウロフが何とかして倍数体に妊性をもたせ,両性生殖を行うクローンを得たいと頑張り続けた。そして約30年を要して異質四倍体の合成に成功した。その研究のあらましはつぎのとおりである。
 |
①
|
二倍体単為発生の誘発=前述の方法で,二倍性メスの卵巣卵を温度処理して,単為発生を誘起させた。単為発生をしたメスの中には,普通の大きさの卵に混じって1.5倍ほどの大きさの大形卵を産む個体があった。また大型卵ばかりを産むメスも少数ながら出現した。おの大型卵については細胞学的ならびに遺伝学的に四倍体であることが確かめられた。
|
| |
② |
この四倍体メスの卵巣卵をとりもう一度温度処理して単為発生を行わせ,四倍性メスのクローンを得た。
|
| |
③ |
有性生殖=②で得た四倍体メスに普通の二倍体オスをかけて,三倍体のメスとオスを得たが,これら同士の交配においても,また三倍体メスに普通の二倍体オスを交配した場合も,また三倍体メスに普通の二倍体オスを交配した場合も,ともに生殖力を欠いていた。
|
| |
④ |
三倍体単為発生処理=そのため三倍体メスに再び温度処理を行い,単為発生を誘発して三倍体メスを得た。この三倍体メスは両性生殖による三倍体メスと異なって,これに普通の二倍体オスを交配すると,今度はわずかながら(約0.5%)妊性を示し,これを孵化させたところ,生存力のあるメスとオスとが生じてきた。
|
| |
⑤ |
この単為発生した三倍体メスの場合にも,二倍体メスに単為発生を行わせた場合に体細胞倍加が起こり二倍性と四倍性の混数体が生じたのと同様に,三倍性+六倍性の混数細胞をもつ個体が見いだされた。この六倍性細胞からは三倍性配偶子ができて,二倍体オスとの交配で,四倍体メスと四倍体オスとができてくる。しかしこれらの四倍体では成熟分裂にあたり,四価の染色体対合をしているものが2〜3組観察されて,そのままでは受精率が低くて,うまく系統を維持することができなかった。
|
| |
⑥ |
異質倍数体の作成=ここに得た四倍体の妊性が低い理由は,この四倍体がカイコ( Bombyx mori , L )起源の染色体だけから構成されている同質四倍体であるため,避けることのできない問題であると考えられたので,アスタウロフはカイコの近縁種のクワコ( Bombyx mandarina )の一系統(シベリアのウスリ地方から上海あたりにかけて生息するクワコで n は28のもの)をかけ合わせて,カイコ 2n ( DD ) + クワコ 2n ( UU )という異質のゲノムから構成される四倍体( DDUU )を作成することを計画した。
そのためにカイコ 4n メスにクワコ 2n オスをかけ合わせ, 3n をつくり,以後前記④,⑤に記した手順をくり返して,ようやくカイコとクワコのゲノム二対ずつをもった異質四倍体(異なったゲノムで構成される倍数体)を合成することができた。この子孫では毎代平均1〜5%程度の妊性を示した。
妊性が低いのは,カイコとクワコとは自然界で交雑が行われるほど近縁なので,両種のゲノムが必ずしも充分に異質でないためと思われる。理想的に隔離されたゲノムを利用して,異質倍数体をつくることができれば,妊性はもっと向上するだろうとアスタウロフは記している。
このような異質倍数体は植物では多数知られ,稔性も正常のものが多い。しかし動物ではまったく珍しい例で,実験的に成功したのはこの例が最初と思われる。
|
|